Коберська В.А., Паламарчук П.П.
Вінницький національний аграрний університет, Україна
МОЛЕКУЛЯРНІ МЕХАНІЗМИ ПРОЯВУ ТОКСИЧНОСТІ МОНОНАТРІЙ ГЛУТАМАТУ
Серед харчових добавок особливе місце належить п ід силювачам смаку (аромату), які використовуються для посилення і/або модифікації натурального смаку і/або аромату харчових продуктів. В чистому вигляді ці речовини не мають ні смаку, ні запаху, але посилюють смак різних страв. В наш час у всьому світі самий відомий посилювач смаку, аромату та замінник солі – мононатрію глутамат Е 621 – натрієва сіль глутамінової амінокислоти (синоніми: глутамат натрію, глютамінат натрію, глутамінат натрію; англ. Monosodium glutamate, скорочено MSG).
В Україні мононатрій глутамат (Е621), так само як і глутамат калію (Е622), кальцію (Е623), амонію (Е624), магнію (Е625), а також сама глутамінова кислота (Е620) дозволені до застосування – до 10 грамів речовини на кілограм продукту, а в приправах і прянощах допустима концентрація ще вище [7 ].
На сьогодні важко знайти консерви, напівфабрикати або готові продукти, виготовлені промисловим шляхом, в яких би не було мононатрій глутамату. У ресторанах швидкого харчування без посилювача смаку не обходиться ні один рецепт. Наприклад, суміші прянощів, що продаються в торгівельних мережах, на третину складаються із глутамату натрію.
Слід відмітити, що в нинішній час рівень додавання глутамату в продукти харчування в 50 разів більший, ніж десятки років тому, і ця цифра продовжує збільшуватись (величина щорічного приросту виробництва нині складає 4%). Щорічна світова потреба в цьому продукті за даними лондонської консалтингової асоціації Л. Гепнера (L. Hepner & Associates) становить близько 1,1 млн. тонн. Зараз глутамат натрію, як харчова приправа, за обсягами виробництва поступається лише солі і перцю.
При цьому допустимі норми можуть бути значно перевищені, що призводить до таких реакцій організму, як головні болі, болі в грудях, часте серцебиття, почервоніння обличчя, приступи гніву та коливання настрою, напади занепокоєння, проблеми поведінки (особливо у дітей та підлітків), спрага, холодний піт, підвищення температури, запаморочення, нудота, розлад шлунку, м’язова слабкість, висипання на шкірі, алергічні реакції, проблеми з диханням, проблеми координації, тощо. Лікарі такі захворювання об'єднали терміном "синдром китайського ресторану" [7; 12; 13]. Кількість добавки, що викликає їх, індивідуальна для кожного, а сама реакція може початися як миттєво, так і протягом 48 годин. Окрім алергічних реакцій можливі набагато серйозніші наслідки регулярного вживання глутамату натрію, що призводить до порушень мозкового кровообігу та, навіть, до хвороб Альцгеймера та Паркінсона. Так, молекули глутамату, здатні зв’язуватись із клітинами сітківки ока і знищувати їх, провокуючи глаукому, яка, переважно, не супроводжується підвищенням внутріочного тиску. Вживання технічного, недостатньо очищеного глутамату натрію , викликає яскраво виражені симптоми , загострення хвороб і навіть – анафілактичний шок, оскільки патогенні домішки синтетичного глутамату діють безпосередньо на кору головного мозку. У поєднанні з вітаміном Е глутамат натрію викликає оксидативне пошкодження клітин організму , при цьому пошкоджуючи ДНК.
Так, як мононатрій глутамат використовується в більшості сучасних харчових технологій в якості харчової добавки і кількість його вживання практично неконтрольований процес, це обумовлює більш детальне вивчення впливу даної харчової добавки на організм людини, перш за все на секреторну функцію шлунку, оскільки шлунок є найбільш уразливим відділом травного тракту, в який потрапляє їжа багата на глутамат, та функціональний стан нервової та імунної системи. Тому метою даної роботи було вивчення механізму впливу мононатрій глутамату на клітинний метаболізм при його надмірному надходженні.
У нормальних умовах глутамат, будучи природнім метаболітом організму, бере активну участь в енергетичному, білковому, жировому обміні (рис. 1).
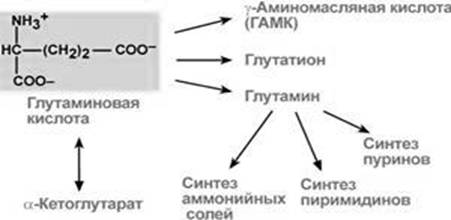
Рис. 1. Шляхи використання глутамату
Незамінна його роль у знешкодженні токсичного аміаку в тканинах. Так, синтез глутаміну – взаємодія глутамату з аміаком – це ключова реакція, що найактивніше відбувається в нервовій та м’язовій тканинах, в нирках, сітківці ока та печінці. Реакція проходить в мітохондріях (рис. 2).
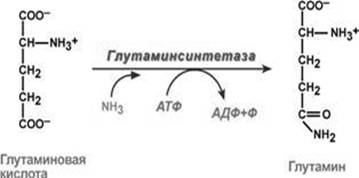
Рис. 2. Взаємодія глутамату з аміаком
Нітроген, що переноситься глутаміном, використовується клітинами для синтезу пуринових та піримідинових кілець, гуанозинмонофосфату (ГМФ), аспарагіну, глюкозаміно-6-фосфату.
Ще значніше прояв цього універсального метаболіту проявляється через стимуляцію гіпофізо-наднирникової осі ендокринної системи. Мобілізація гормонів надниркової залози забезпечує підвищення стійкості до гіпоксичних, фізичних, психоемоційних перевантаженням, знижує запальні, алергічні прояви [4].
Глутамат синтезується в нейронах зі свого попередника глутаміну, накопичується в синаптичних везикулах і в підсумку вивільняється за допомогою кальцій залежного механізму. Глутамат, який вивільнився може взаємодіяти із глутаматними рецепторами будь-якого типу, що перебувають у синаптичній щілині. В астроцитах глутамат захоплюється й перетворюється в глутамін за допомогою глутамінсинтетази.
Для нормального протікання синтезу глутамату в організмі необхідна наявність у субстраті неорганічного фосфату, а також макро- і мікроелементів, що входять до складу ферментних систем даного процесу. Синтез глутаміну відбувається згідно схеми, наведеної на рис. 3.
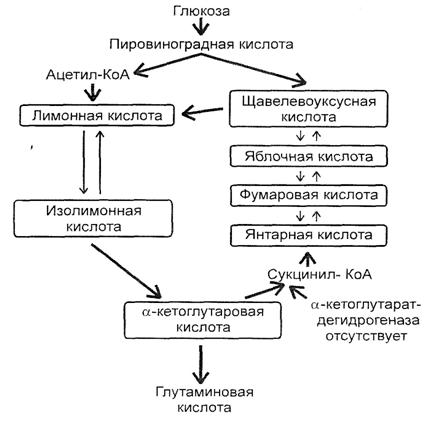
Рис. 3. Синтез глутамінової кислоти
Глутамат натрію, сам по собі не має ні смаку, ні запаху, хоча володіє властивістю підсилювати смак харчового продукту. При потраплянні в організм він легко проникає в кров, мозок і напряму впливає на мутацію генів, що відповідають за смакові відчуття, при чому посилюється чутливість смакових сосочків язика. Варто зазначити, що глутамат натрію діє за принципом наркотику, провокуючи поступове звикання, це призводить до втрати чутливості смакових рецепторів. На сьогодні добре відомо, що глутамат є головним збудливим нейротрансмітером в центральній нервовій системі ссавців [8; 10], який реалізує свою дію через метаботропні (mGluR 1 – mGluR 8 ) та іонотропні (НМДА-, каїнатний і АМПА- підтипи) глутаматні рецептори (іГлуР). Глутаматні механізми представлені приблизно у 40% нервових клітин, а частина, що залишилася припадає на частку всіх інших медіаторів (серотоніну, ацетилхоліну, дофаміну та ін.).
Іонотропні рецептори, з'єднані з іонними каналами, вони відкривають їх після активації відповідними молекулами (лігандами), так що потоки іонів викликають електричну активність нейрону. Метаботропні рецептори структурно не пов'язані з іонними каналами, вони управляють метаболічними процесами в клітині через спеціальні сигнальні молекули-інформатори, контролюючи активність іонотропних рецепторів. Ліганди, що активують нейрональні рецептори, – їх первинні інформатори (первинні месенджери), а сигнальні молекули, які утворюються при активації метаботропних рецепторів і використовуються для корегування сигналів усередині клітини, – вторинні месенджери. Загалом, порушення в клітині, викликані іонотропними рецепторами синаптичного контакту, контролюються метаботропними рецепторами цих же синаптичних мембран (рис. 4).
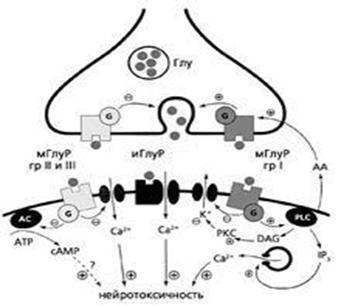
Рис. 4. Схема взаємодії іонотропного і метаботропних рецепторів нейрона
В організмі глутамат натрію звільняється від іона натрію, перетворюючись у глутамінову кислоту, або глутамат. Останній у тканинах головного мозку послідовно перетворюється в протектор нейронів головного мозку, надалі в бурштинову кислоту, найбільш ефективний енергетичний субстрат. При ускладненому статусі, глутамат, через глутаматзалежні рецептори нейрона, запускає глутамат-кальцієвий каскад реакцій, який веде до перезбудження і виснаження клітини [2]. Дослідження причин глутаматної нейротоксичності показали, що вхід Са 2+ через НМДА-підтип глутаматних каналів і послідуюче накопичення Са 2+ в мітохондріях являються основними факторами, що призводять до загибелі клітин. Таким чином, взаємодія між каїнатним і НМДА-рецепторами визначає тривалість хвилі збудження і ефективність перебудови метаболізму нервової клітини під впливом кальцію.
За даними Burns G.A., Stephens K.E., 1995 [6], в ентеральних нейронах шлунково-кишкового тракту знайдена мРНК, яка кодує НМДА-підтип глутаматних рецепторів. Також фармакологічні дослідження показали, що у регуляції моторної [11] та секреторної [9] функцій кишечника беруть участь глутаматні рецептори.
Так, щоденне введення мононатрійглутамату натрію щурам у дозі 15 та 30 мг/кг (1 та 2 грами на середньостатистичну людину) впродовж 20-ти та 30-ти днів викликає гіперсекрецію хлоридної кислоти, що може бути причиною патогенезу кислотозалежних захворювань. Отже, крім основної ролі збудливого нейромедіатора, глутамат може проявляти нейротоксичні властивості. При гіперактивації глутаматергічної передачі відбувається інтенсивне надходження іонів кальцію в клітину. Підвищений вміст вільного кальцію здатний продукувати процеси утворення реактивних форм кисню, наслідком чого може бути ушкодження й загибель клітин.
Відомі факти, що вказують на можливу присутність глутаматних рецепторів не тільки в нейрональних клітинах [1]. Виявлено, що глутамат добре зв'язується з мембранами лімфоцитів людини [3]. Пізніше було показано, що глутаматні рецептори є у лімфоцитах і їх активація призводить до зростання в клітинах вільних іонів кальцію і активних форм кисню, в результаті чого активується каспаз 3, що відкриває клітині шлях до апоптозу [5]. Навіть після короткої інкубації з'являлася велика кількість мертвих клітин. Отже, можна сказати, що присутність іонотропного і метаботропних рецепторів глутамату на мембранах лімфоцитів робить їх чутливими до тих же сигнальних молекул, які управляють активністю нейронів (рис. 5).
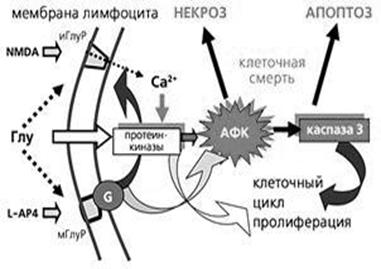
Рис. 5. Регулювання життя і смерті лімфоцита глутаматними рецепторами
При взаємодії глутамату (Глу) з іонотропними рецепторами (іГлуР) іони кальцію входять всередину клітини, активують протеїнкінази і каспаз 3, яка стимулює проліферацію. Взаємодія глутамату з метаботропними рецепторами (мГлуР) через G-білки стимулює активність іГлуР, що призводить до додаткової активації протеїнкінази і посиленого росту активних форм кисню (АФК). У цьому випадку можлива індукція клітинної смерті як по шляху апоптозу, так і по дорозі некрозу. Наявність глутаматних рецепторів у клітинах імунної системи розкриває структурну основу цих взаємодій і дозволяє вважати глутамат не тільки нейро-, але і імуномедіатором.
Висновок. Отже, максимальні добові дози даної харчової добавки мають бути переглянуті з урахуванням їх нейротоксичності, негативного впливу на діяльність травної системи, на процеси утворення активних форм кисню, що може провокувати переродження та загибель клітин організму.
Список використаних джерел:
1. Болдирєв А.А., Тунева Є.О. // Біологічні мембрани. – 2005. – Т. 22. – С.142–145.
2. Копаниця М.В. Позасинаптичні рецептори нейротрансмітерів: поширення, механізми активації та фізіологічна роль / М.В. Копаниця // Нейрофизиология. – 1997. – 27. – 6. – С. 448–458.
3. Костанян І.А., Наволоцький Є.В., Нурієв Р.І. та ін. // Біоорганічна хімія. – 1997. – Т. 23. – С. 805–808.
4. Раевский К.С., Георгиев В.П. Медиаторные аминокислоты: нейрофармакологические и нейрохимические аспекты / К.С. Раевский, В.П. Георгиев. – М.: Медицина, 1986. – 239 с.
5. Boldyrev A.A., Kazey V.I., Leinsoo T.A. et al. // Biochem. Biophys. Res. Commun. 2004. – V. 324. – P.133–139.
6. Burns G.A., Stephens K.E. Expression of mRNA for the N-methyl-D-aspartate (NMDAR1) receptor and vasoactive intestinal polypeptide (VIP) co-exist in enteric neurons of the rat // J. Auton. Nerv. Syst. – 1995. – Vol. 55. – P. 207–210.
7. Freeman M. Reconsidering the effects of monosodium glutamate: a literature review / M. Freeman // J Am Acad Nurse Pract. 2006. – Vol. 18, № 10. – P. 482–486.
8. Gasic G. System and moleculargenetic approaches converge to tackle learning and memory // Neuron. – 1995. – Vol. 15 – P. 507–512.
9. Rhoads J.M., Argenzio R.A., Chen W., Gomez G.G. Asparagine stimulates piglet intestinal Cl secretion by a mechanism requiring a submucosal glutamate receptor and nitric oxide // J. Pharmacol. Exp. Ther. – 1995.- Vol. 274. – P. 404–412.
10. Watkins J.C., Jane D.E. The glutamate story // Br. J. Pharmacol. – 2006. – V. 147. – P. 100–108.
11. Wiley J.W., Lu Y., Owyang C. Evidence for a glutamatergic neural pathway in the myenteric plexus // Am. J. Physiol. – 1991. – Vol. 261. – P. 693–700.
12. Williams A.N. Monosodium glutamate 'allergy': menace or myth? / A.N. Williams, K.M. Woessner // Clin Exp Allergy. – 2009. – 39, № 5. – P. 640–646.
13. Yang W.H. The monosodium glutamate symptom complex: assessment in a double-blind, placebo-controlled, randomized study / W.H. Yang, M.A. Drouin, M. Herbert et al. // J Allergy Clin Immunol. – 1997. – 99. – P. 757–762.